Классификация ядерных рецепторов и их лиганды
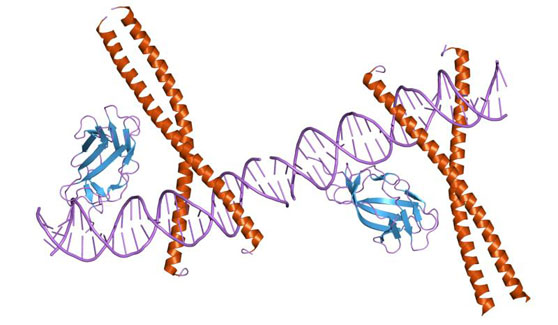
Домены ядерных рецепторов
Ядерные Рц имеют модульную (доменную) организацию, включающую за редкими исключениями 4—5 независимых, но функционально связанных доменов.
Модуляторный домен А/В наиболее вариабелен как по длине, так и по последовательности аминокислот. Часто варианты Рц возникают за счёт модификаций именно этого домена. Локализованная в этом домене автономная транскрипционная активационная функция 1 (AF-1) действует синергично с лигандзависимой активационной функцией 2, локализованной на противоположном конце молекулы Рц, и обеспечивает специфичность активности Рц в соответствии с контекстом контролируемого промотора и данного типа клеток, что свидетельствует о взаимодействии этого домена с тканеспе-цифичными кофакторами. В некоторых Рц (например, Рц В прогестерона) выделяют ещё одну активационную функцию (AF-3), также локализованную в домене А/В. Различия в структуре домена А/В могут обеспечивать различия в действии одного и того же лиганда на активность родственных Рц. Домен А/В служит мишенью для действия ряда протеинкиназ, модулирующих активность Рц (см. ниже). Показано прямое взаимодействие этого домена с группоспецифичными (SRCs, рЗОО, СВР) и избирательными (MMS19, р72/р68 белки с доменом DEAD) коактиваторами ядерных Рц, ведущее к повышению активности Рц.
ДНК-связывающий домен и гормончувствительные элементы.
ДНК-связывающий домен (домен С) наиболее консервативен. Он узнает энхансерные участки ДНК, называемые гормончувствительными элементами (HREs), которые состоят из одного или двух консенсусных полусайтов, распознаваемых соответственно моно- и димерными Рц.
Сдвоенные полусайты
Сдвоенные полусайты могут быть организованы как палиндромы (т.е. структуры с комплементарной симметрией) и прямые и обратные повторы. Рц стероидов (исключая Рц эстрогенов) узнают консенсусный полусайт AGAACA, а Рц эстрогенов и другие ядерные Рц — полусайт AGGTCA. HREs для мономерных Рц содержат 5′-расширение из 3-6 оснований, обогащенное А/Т. Природные HREs редко полностью соответствуют указанным консенсусным последовательностям. В гене может содержаться один или несколько HRE для данного гормона, локализованных, как правило, в 5′-примыкающей области, и реже — в последовательностях интронов, экзонов и З’-примыкающей области. Множественность HRE обеспечивает кооперативное усиление действия лиганда.
Осуществлён рентгеноструктурный анализ изолированных ДНК-связывающих доменов и их димеров для ряда Рц, позволивший получить детальную картину Рц-нуклеинового взаимодействия. ДНК-связывающий домен состоит их трёх модулей: двух «цинковых пальцев» (суммарно 66—70 а.к. остатков) и С-концевого расширения (около 25 остатков).
Аминокислотные остатки первого цинкового пальца (область Р-Ьох) непосредственно узнают полусайт HRE. Второй цинковый палец взаимодействует с сахарофосфатным остовом ДНК неспецифично и служит одной из поверхностей димеризации Рц (области D- и DR-блоков). С-концевое расширение выполняет несколько функций: взаимодействует с прилегающими к полусайту HRE остатками оснований (что особенно важно для специфичности и прочности связывания с HRE мономерных Рц и для правильной ориентации гетеродимерных Рц, взаимодействующих с несимметричными HPvEs), создаёт поверхность взаимодействия Рц с другими белками и служит молекулярной линейкой для дифференцировки HREs по размеру вставки (спейсера), разделяющей два полусайта HRE. О Размер вставки (спейсера), разделяющей два полусайта HRE, может варьировать от 0 до 5-6 пар нуклеотидов (п.н.). В случае HREs для Рц стероидов спейсер состоит из 3 п.н.
Шарнирная область
Другие ядерные Рц нередко могут узнавать более одного типа HREs в зависимости от партнёра по димеризации. В случае HREs, организованных в виде прямых повторов, размер спейсера играет важную роль в избирательности взаимодействия HRE с разными Рц.
- Шарнирная область D Рц вариабельна по размеру и аминокислотной последовательности. Её гибкость способна обеспечить поворот ДНК-связывающего домена на 180° относительно лигандсвязывающего домена для создания возможности взаимодействия димерных Рц с несимметричными HREs, представленными прямыми повторами, и с HREs, представленными инвертированными повторами. Эта область также создаёт поверхность взаимодействия Рц с некоторыми корегуляторами и может содержать сигнал ядерной локализации.
- Лигандсвязывающий домен Е умеренно консервативен. В нём содержится общий для ядерных Рц мотив (сигнатура) в области четвёртой спирали и высококонсервативная последовательность, локализованная на С-конце молекулы (во многих Рц 12-я спираль), соответствующая зависимой от лиганда активационной функции AF-2. Рентгеноструктурный анализ лигандсвязывающего домена многих Рц в форме апобелков или комплексов с лигандами показал консервативность общего плана строения этого домена: 11—13 а-спиралей уложены в три слоя, в результате чего гидрофобный лигандсвязывающий «карман» оказывается в глубине лигандсвязывающего домена. Помимо связывания лиганда, этот домен выполняет ряд других функций: участвует в димеризации Рц, взаимодействует с белками теплового шока, содержит сигнал ядерной локализации, участвует в трансактивационном действии Рц. Последняя функция осуществляется при обязательном участии AF-2 посредством рекрутирования коактиваторов.
Конформационное изменение Рц
Механизм работает по типу мышеловки: связывание лиганда вызывает конформационное изменение Рц, заключающееся в изменении ориентации AF-2-мотива, в результате чего лиганд оказывается «запертым», и на месте входа в лигандсвязывающий карман возникает поверхность (включающая AF-2 и ещё по меньшей мере 3 спирали) взаимодействия с коактиваторами. Антагонисты Рц не способны индуцировать подобные изменения конформации Рц и соответственно обеспечивать его взаимодействие с коактиваторами. Селективные агонисты/антагонисты, называемые также селективными модуляторами Рц, индуцируют «неправильную» ориентацию Н12, что обеспечивает активность Рц только в определённом генном и клеточном контексте. Данное направление в фармакологии представляется чрезвычайно перспективным, и знание структуры лигандсвязывающих доменов ядерных Рц создаёт реальную возможность конструирования необходимых клиницистам лигандов с заданными биологическими свойствами.
Домен F, имеющийся не во всех ядерных Рц, может обеспечивать дополнительную дискриминацию агонистов и антагонистов Рц. Например, его удаление из Рц прогестерона или глюкокортикоидов превращает антагонисты Рц в агонисты. Предполагают, что этот домен может быть связан с неким корепрессором.
Функционально домены тесно связаны между собой, что, в частности, находит отражение в изменениях способности отдалённых доменов Рц взаимодействовать с другими молекулами в результате связывания лиганда, ДНК и других молекул за счёт конформационной пластичности Рц.
Экспрессия ядерных рецепторов
Уровень ядерных Рц служит объектом контроля со стороны множества факторов, включая собственные лиганды этих Рц: в генах многих ядерных Рц обнаружены гормончувствительные элементы, обеспечивающие систему (положительной или отрицательной) обратной связи. Экспрессия Рц может быть ограничена одной или немногими тканями [например, в случаях LXR (преимущественно в печени) или стероидогенного фактора 1 SF-1 (в гонадах, надпочечниках, гипофизе, гипоталамусе) или быть практически повсеместной (например, RXR)]. Включение экспрессии может происходить на самых ранних этапах эмбрионального развития (например, SF-1, определяющего развитие гонад) или на более поздних этапах (например, Рц андрогенов и эстрогенов, обеспечивающих действие продуктов сформированных и функционирующих гонад — андрогенов и эстрогенов). Как правило, экспрессия ядерных Рц сохраняется в ходе дальнейшего индивидуального развития, но может снижаться при старении. Уровень ядерных Рц в значительной мере определяет чувствительность клеток к гормонам, что, в частности, давно нашло применение в клинике для прогнозирования эффективности гормонотерапии.
Внутриклеточная локализация
Большинство ядерных Рц конститутивно (т.е. независимо от наличия лиганда) локализовано почти исключительно в клеточном ядре, тогда как значительная или основная часть Рц стероидов в отсутствие лиганда может находиться в цитоплазме. Удержание Рц в клеточном ядре, по-видимому, в значительной мере определяется межбелковыми взаимодействиями (например, с партнёром по димеризации PvXR и корепрессором NCoR), а не связыванием с ДНК. Независимо от типа Рц, соответствующий лиганд вызывает внутриядерное перераспределение Рц между нуклеоплазмой и хроматином, гетеро- и эухроматином. Указанная выше особенность Рц стероидов обусловлена их способностью связываться в цитоплазме с белками теплового шока (heat shock proteins — Hsp), которые препятствуют транспорту Рц через ядерную мембрану (осуществляемому благодаря наличию в лигандсвязывающем домене Рц последовательности, служащей сигналом ядерной локализации) и связыванию Рц с ДНК (путём экранирования ДНК-связывающих доменов Рц).
Связывание лиганда
Зрелая форма Рц стероидов — комплекс Рц с димерным Hsp90, включающий также иммунофилин и шаперонный белок р23. Процессинг новосинтезированного Рц стероидов может включать АТР-зависимую и независимую ассоциацию Рц с, по меньшей мере, 8 шаперонными белками, значение которой, по-видимому, заключается в последовательном придании полипептидной цепи Рц необходимой конформации.
В отсутствие лиганда ядерные Рц могут быть транскрипционно неактивными (например, Рц стероидов), оказывать репрессорное (например, Рц тиреоидных гормонов) или стимулирующее (например, конститутивно активные Рц CAR) действие на транскрипцию. Связывание лиганда ведёт к изменению транскрипционной активности Рц, индуцируя, усиливая или ослабляя исходную активность Рц. Эти изменения опосредованы взаимодействием Рц с корегуляторными белками. В зависимости от состояния Рц эти корегуляторы, во-первых, создают транскрипционно пермиссивную или не-пермиссивную среду в области промотора контролируемого гена и, во-вторых, связывают Рц с общими транскрипционными факторами и РНК-полимеразой II. Установлено и прямое (зависимое от лиганда) связывание ряда ядерных Рц с отдельными компонентами транскрипционного комплекса (например, с ТВР, TFIIF, TFIIH и TFIIB). Предполагают, что такое взаимодействие может влиять на сборку транскрипционных комплексов. Однако избирательность эффектов ядерных Рц (зависимость от типа клетки и контролируемого промотора) достигается, по-видимому, в значительной мере за счёт их регулируемого лигандом взаимодействия с корепрессорами, коактиваторами и коинтеграторами СВР (CREB-связывающим белком)/р300.
Коактиваторы. Известно несколько типов коактиваторов ядерных Рц. Структурно-функциональная организация членов семейства коактиваторов Рц стероидных гормонов (SRC) представлена на примере коактиватора SRC-1.
- Предыдущие статьи из рубрики: Эндокринология
 Общие свойства рецепторов
Общие свойства рецепторовРецепция и проведение сигнала Эти системы могут включать так называемые «вторые посредники» — низкомолекулярные соединения (циклические нуклеотиды, фосфолипиды […]
 Функции транспортных белков
Функции транспортных белковАнгаотензинсвязывающий белок Ангаотензинсвязывающий белок, возможно, появляется в крови при патологии и идентичен митохондриальной олигопептидазе М; белок, связывающий адреномедуллин, […]
 Транспорт сигнальных соединений
Транспорт сигнальных соединенийТранспортные белки и их лиганды К известным транспортным белкам относятся белки, связывающие стероиды (1), тиреоидные гормоны и ретиноиды […]
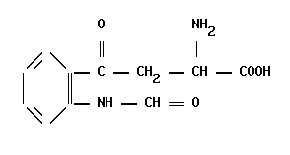 Производные триптофана
Производные триптофанаБелково-пептидные сигнальные соединения Ацетилхолин синтезируется в синапсах из холина и ацетил-КоА под действием холин-О-ацетилтрансферазы. В отличие от низкомолекулярных […]
 Тиреоидные гормоны
Тиреоидные гормоныЙодирование тироглобулина происходит в комплексе Гольджи у апикальной поверхности клеток под действием тиропероксидазы — гемсодержащего белка, катализирующего 1) […]