Общие свойства рецепторов
Рецепция и проведение сигнала
 Эти системы могут включать так называемые «вторые посредники» — низкомолекулярные соединения (циклические нуклеотиды, фосфолипиды и их метаболиты, ионы кальция), а также разнообразные ферменты, адапторные белки, транскрипционные факторы, активность которых регулируется непосредственно Рц, вторыми посредниками или зависимыми от сигнальных соединений белками-участниками каскадов. Несмотря на разнообразие систем проведения сигнала, многие элементы этих систем оказываются общими в действии не только родственных, но и химически далёких друг от друга сигнальных соединений, действующих через Рц разных классов. В результате такой «вырожденности» путей проведения сигнала ряд конечных физиологических эффектов разных сигнальных соединений оказывается сходным. В качестве примера можно привести гипергликемию, индуцируемую гормонами стресса (катехоламинами, глюкокортикоидами, гормоном роста). Вместе с тем, за редкими исключениями спектр индуцируемых в клетке изменений для каждого сигнального соединения индивидуален, что определяет качественное своеобразие эффектов данного регулятора. При совместном действии на клетку нескольких сигнальных соединений, что является нормальным состоянием клетки, индивидуальные системы проведения сигнала взаимодействуют друг с другом, приводя в конечном итоге к сложнейшей мозаике взаимного усиления одних эффектов, ослабления других и независимости третьих.
Эти системы могут включать так называемые «вторые посредники» — низкомолекулярные соединения (циклические нуклеотиды, фосфолипиды и их метаболиты, ионы кальция), а также разнообразные ферменты, адапторные белки, транскрипционные факторы, активность которых регулируется непосредственно Рц, вторыми посредниками или зависимыми от сигнальных соединений белками-участниками каскадов. Несмотря на разнообразие систем проведения сигнала, многие элементы этих систем оказываются общими в действии не только родственных, но и химически далёких друг от друга сигнальных соединений, действующих через Рц разных классов. В результате такой «вырожденности» путей проведения сигнала ряд конечных физиологических эффектов разных сигнальных соединений оказывается сходным. В качестве примера можно привести гипергликемию, индуцируемую гормонами стресса (катехоламинами, глюкокортикоидами, гормоном роста). Вместе с тем, за редкими исключениями спектр индуцируемых в клетке изменений для каждого сигнального соединения индивидуален, что определяет качественное своеобразие эффектов данного регулятора. При совместном действии на клетку нескольких сигнальных соединений, что является нормальным состоянием клетки, индивидуальные системы проведения сигнала взаимодействуют друг с другом, приводя в конечном итоге к сложнейшей мозаике взаимного усиления одних эффектов, ослабления других и независимости третьих.
Общие свойства Рц: связывание лигандов, конформационные изменения, олигомеризация и десенситизация.
Лигандсвязывающие свойства
Общим для всех Рц является их способность избирательно и с высоким сродством связывать природные сигнальные соединения, их синтетические аналоги и антагонисты. Равновесная константа взаимодействия может варьировать в весьма широком диапазоне (Ю-6 >Kd >10″п М) в соответствии с физиологическим уровнем данного сигнального соединения. Например, глюкокортикоиды, концентрация которых в крови составляет приблизительно Ю-7 М, связываются со своим Рц с Kd ~10~7—10~8 М (при О °С), а эстрогены, уровень которых в крови на 3 порядка ниже уровня глюкокортикоидов, связываются со своим Рц с К, ~1010 М. Кинетика взаимодействия также широко варьирует. Как правило, чем ниже сродство Рц к лиганду, тем быстрее происходят процессы ассоциации и диссоциации. Так, при физиологических условиях ассоциация нейромедиаторов с Рц занимает доли секунды, а для насыщения стероидными гормонами Рц требуются многие минуты. В пределах группы родственных соединений, взаимодействующих с одним Рц, величина сродства определяется в первую очередь скоростью диссоциации лиганд—рецепторных комплексов.
Сродство Рц к лиганду может быть функцией времени. В случае белковых лигандов их взаимодействие с Рц может происходить по принципу застёжки-«молнии», когда взаимодействие первых фрагментов Рц и лиганда облегчает последующее взаимодействие молекул по другим пунктам. Другим вариантом изменения сродства во времени служит индуцированная лигандом сборка олигомерного Рц. Благодаря появлению новых точек контакта между лигандом и Рц такой олигомер связывает лиганд значительно прочнее, чем составляющие его СЕ в отдельности.
Величина сродства Рц к лиганду может служить объектом регуляции, включая ауторегуляцию. Наиболее известным вариантом такой регуляции является изменение характера фосфорилирования Рц, сопровождающееся снижением сродства в результате изменения конформации Рц или присоединения к нему регуляторного белка. Выявлены также регуляторные белки, изменяющие соотношение сродства Рц к родственным лигандам.
Конформационные изменения
Связывание сигнального соединения с Рц вызывает изменения в конформации последнего, сопровождающиеся изменением способности Рц взаимодействовать с нижележащими элементами системы проведения сигнала (эффекторными молекулами) или собственной ферментативной активности Рц. Вариантом таких изменений служит индукция лигандом сборки (или диссоциации на СЕ) олигомерного Рц.
Существуют две основные концепции, объясняющие механизм индуцируемого лигандом конформационного перехода Рц. Первая из них, основывающаяся на представлениях классической энзимо-логии и широко признаваемая специалистами в области Рц, сопряжённых с G-белками, постулирует существование двух конформа-ционных состояний Рц, активного и неактивного. Связывание агониста стабилизирует активную конформацию Рц, что вызывает смещение равновесия между активной и неактивной конформациями в сторону первой. Антагонист оказывает противоположное действие. Согласно другой концепции, принимаемой специалистами в области ядерных Рц, лиганд активно изменяет конформацию Рц. При этом допускается существование множества конформационных состояний Рц. Достоинством последней модели, в частности, является возможность объяснения существования частичных или избирательных агонистов/антагонистов, воспроизводящих одни эффекты полного агониста и не воспроизводящих другие его эффекты.
Олигомеризация
Рц ряда классов олигомеризуются под действием лиганда. В ряде случаев эта олигомеризация является необходимым этапом в действии сигнальных соединений. Конкретная функция олигомеризации различна для разных Рц. В случае Рц-тирозинкиназ и серии/ треонинкиназ олигомеризация обеспечивает сближение фермента (внутриклеточного домена одной СЕ Рц) с субстратом (внутриклеточным доменом другой СЕ Рц).
Механизмы самоограничения
В случае ядерных Рц димеризация способствует стабилизации связывания Рц с ДНК и корегуляторами. Вместе с тем известны примеры действия ядерных Рц в форме мономеров. Рц-протеинфосфатазы, напротив, активны лишь в форме мономеров, но не димеров. Рц-гуанилатциклазы — конститутивные димеры. В случае Рц, сопряжённых с G-белками, обнаружена как гомо-, так и гетероолигомеризация, которая может позитивно или негативно влиять на проведение сигнала и десенситизацию Рц. Таким образом, индуцируемая лигандом олигомеризация Рц является необходимым элементом проведения сигнала в одних случаях, факультативным элементом в других, отсутствующим элементом в третьих и блокирующим элементом в четвертых.
Системы рецепции включают механизмы самоограничения развития эффектов; часть этих механизмов связана с регуляцией количества и/или функциональной активности Рц. В случае мембранных Рц одним из общих механизмов служит индуцируемая сигнальным соединением кластеризация (образование скоплений) Рц, их последующая интернализация (захват внутрь клетки) и частичная или полная деградация с помощью ряда систем протеолиза. С данным механизмом тесно связаны зависимые от лиганда процессы фосфорилирования/дефосфорилирования Рц, которые могут играть и самостоятельную роль в десенситизации. В случае ядерных Рц десенситизация в ряде случаев также связана с изменениями в уровне фосфорилирования. Кроме того, индуцируемые лигандом изменения конформации Рц способствуют вступлению Рц на путь деградации. Важно отметить, что десенситизация тесно связана с механизмами активации Рц.
Рецепция сигнальных соединений имеет важнейшее значение для клинической медицины. Многие лекарственные препараты представляют собой аналоги (агонисты или антагонисты) природных сигнальных соединений. Развитие ряда заболеваний связано с мутациями генов, кодирующих рецепторные белки и другие элементы системы проведения сигнала. Эпидемиологические исследования показывают, что имеющийся в популяции полиморфизм генов Рц, не затрагивающий функциональные свойства белков, также может иметь значение для предрасположенности к различным заболеваниям или ускоренному старению.
Ядерные рецепторы
Ядерные Рц — транскрипционные факторы, которые обеспечивают прямой контроль экспрессии генов со стороны сигналов внешней среды и внутренней среды организма. Активность этих Рц контролируется, по меньшей мере, тремя механизмами: 1) взаимодействием с небольшими липофильными лигандами; 2) ковалентной модификацией, обычно фосфорилированием, регулируемой вне- и внутриклеточными сигналами; 3) межбелковыми взаимодействиями с другими транскрипционными факторами.
К настоящему времени клонировано 49 представителей надсемейства ядерных Рц человека. В зависимости от сходства аминокислотных последовательностей и ряда других свойств эти белки группируют в несколько подсемейств. Классификация ядерных Рц, обнаруженных у человека, представлена в табл. Продукты па-ралогичных (т.е. структурно и функционально близкородственных) генов обозначены греческими буквами а, р, у, 8. Кроме того, многие Рц представлены несколькими вариантами, возникающими в результате использования альтернативных промоторов, альтернативного сплайсинга транскриптов или альтернативных стартов трансляции. Как правило, такие варианты обозначают арабскими цифрами 1, 2 и т.д. Экспрессия разных генов Рц одного типа и их вариантов тканеспецифична и имеет индивидуальные особенности регуляции, в частности в онтогенезе.
Лиганды ядерных Рц могут представлять собой как гормоны, так и негормональные соединения (например, продукты, поступающие с пищей, и их метаболиты, а также продукты метаболизма широко распространённых эндогенных соединений, например холестерина и полиненасыщенных жирных кислот). Ряд лекарственных препаратов и поллютанты также могут выступать в качестве лигандов ядерных Рц.
- Предыдущие статьи из рубрики: Эндокринология
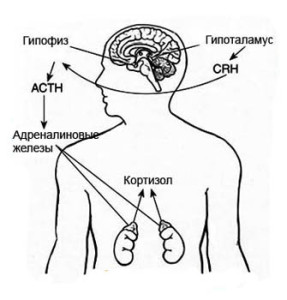
 Функции транспортных белков
Функции транспортных белковАнгаотензинсвязывающий белок Ангаотензинсвязывающий белок, возможно, появляется в крови при патологии и идентичен митохондриальной олигопептидазе М; белок, связывающий адреномедуллин, […]
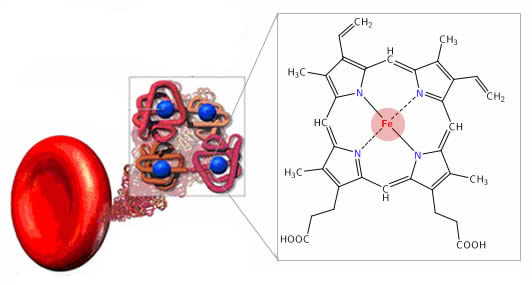
 Транспорт сигнальных соединений
Транспорт сигнальных соединенийТранспортные белки и их лиганды К известным транспортным белкам относятся белки, связывающие стероиды (1), тиреоидные гормоны и ретиноиды […]
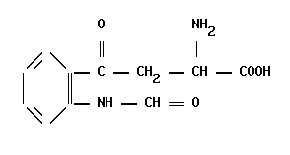 Производные триптофана
Производные триптофанаБелково-пептидные сигнальные соединения Ацетилхолин синтезируется в синапсах из холина и ацетил-КоА под действием холин-О-ацетилтрансферазы. В отличие от низкомолекулярных […]
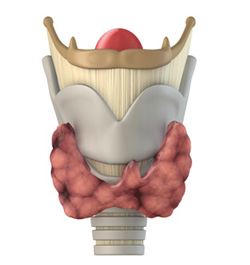 Тиреоидные гормоны
Тиреоидные гормоныЙодирование тироглобулина происходит в комплексе Гольджи у апикальной поверхности клеток под действием тиропероксидазы — гемсодержащего белка, катализирующего 1) […]
 Производные аминокислот
Производные аминокислотФосфолипазы А2 — большое семейство (известно около 20 членов) белков с различающейся субстратной предпочтительностью, локализацией (выделяют секретируемые, зависимые […]