Классификация ядерных рецепторов и их лиганды
Особенности некоторых ядерных рецепторов
Данный факт объясняет эстрогеноподобное действие эпидермального фактора роста (EGF), активирующего МАРК, в матке и опухолях молочной железы. Важно отметить, что фосфорилирование Рц может по-разному сказываться на его активности в разных клетках и на разных промоторах, что, видимо, отражает разнообразие взаимодействующих с Рц корегуляторов.
Помимо фосфорилирования, активность ядерных Рц могут регулировать и другие ковалентные модификации (например, повышение транскрипционной активности Рц андрогенов в результате ацетилирования коактиватором ТірбО и снижение активности после деацетилирования гистондеацетилазой 1; оба белка могут быть ассоциированы с данным Рц). Механизм потенцирующего влияния ацетилирования на активность Рц может быть связан со снижением сродства Рц к корепрессорам.
PPARs. Широкий интерес к Рц активаторов пролиферации пероксисом (PPARs) определяется, во-первых, ключевой ролью этих белков в регуляции энергетического и липидного обменов и, во-вторых, выявлением того факта, что ряд противодиабетических, гиполипидемических и противовоспалительных лекарственных препаратов оказывает свое действие через PPARs. PPARoc интенсивно экспрессируется в сердце, печени, почках, кишечнике, буром жире, т.е. в тканях с высокой скоростью р-окисления жирных кислот. Уровень PPARoc контролируют стрессорные воздействия, глюкокортикоиды, инсулин. PPARp экспрессируется более широко, включая мозг, почки, кишечник, клетки Сертоли. Изоформы PPARy (продукты альтернативного сплайсинга) образуются тканеспецифично: PPARy 1 много в селезёнке, кишечнике, белом жире, a PPARy2 — в белом и буром жире. PPAR-чувствительные элементы (PPREs) — прямые повторы со спейсером из 1 п.н. и 5′-расширением — консенсус 5′-ААСТ AGGNCA A AGGTCA — обнаружены в генах, контролирующих обмен углеводов и липидов. PPARs функционируют в виде гетеродимеров с RXR, причём действие лигандов обоих партнёров аддитивно (например, действие тиазолидиндионов in vivo в снижении гипергликемии и гипертриацилглицеридемии при голодании усиливается одновременным введением 9-г<«с-ретиноевой кислоты).
Катаболизм липидов
Помимо RXR, партнёрами PPAR по гетеродимеризации могут служить TRp и LXRa. PPARa активируется жирными кислотами, эйкозаноидами, карбапростациклином, нестероидными противовоспалительными препаратами, лейкотриеном р4 (LTB4). PPARp и PPARy также активируются общими для всех PPAR лигандами [докозагексеновой кислотой, некоторыми простагландинами (PGs)]. Кроме того, PPARy специфически активируют тиазолидиндионы (группа противодиабетических лекарств), метаболиты PGs 15-де-зокси-Д1214 — PGJ2, полиненасыщенные жирные кислоты и нестероидные противовоспалительные средства (например, ибупрофен). Сравнительно низкая избирательность PPARs в отношении лигандов при сравнительно низком сродстве определяется особенностями лигандсвязывающего кармана этих Рц, который, во-первых, по объёму значительно превышает объём самих лигандов, во-вторых, имеет Т- или Y-образную форму, обеспечивающую адаптацию разных лигандов, и, в-третьих, содержит дополнительный вход. Помимо низкомолекулярных лигандов, регуляторами активности PPARs могут служить протеинкиназы (в частности, МАРК). Корегуляторами транскрипционной активности PPARs служат как общие для ядерных Рц факторы (SRC-1, СВР/рЗОО, SMRT), так и более избирательные коактиваторы PPARy (PGC-1 и PGC-2).
В физиологическом плане PPARa и PPARy являются в целом антагонистами, поскольку первый ускоряет катаболизм липидов, а второй стимулирует их анаболизм. PPARa стимулирует биосинтез белков, обеспечивающих транспорт липидов через энтероциты в лимфу и из крови — в гепатоциты. В печени PPARa стимулирует утилизацию жирных кислот посредством усиления экспрессии ацил-КоА-синтазы, ацил-КоА-оксидазы, бифункционального фермента, кетоацил-КоА-тиолазы. После нескольких циклов укорочения жирных кислот при р-окислении в пероксисомах образовавшиеся продукты направляются в митохондрии для дальнейшей деградации, которую также стимулирует PPARa за счёт индукции карнитин-пальмитоилтрансферазы и ацил-КоА-дегидрогеназы для цепей среднего размера. Один из путей ускорения утилизации жирных кислот под действием PPARa — индукция митохондриальной 3-гидрокси-З-метилглутарил-КоА-синтазы, обеспечивающей образование кетоновых тел.
Деградации жирных кислот в печени
Деградации жирных кислот в печени способствует также торможение под действием PPARa биосинтеза ряда аполипопротеинов, предназначенных для экспорта триглицеридов из печени. Голодание и стресс оказывают во многом сходное действие на обмен веществ благодаря PPARoc: при голодании из жировых депо высвобождаются жирные кислоты, являющиеся активаторами PPARa, а глюкокортикоиды при стрессе являются индукторами экспрессии PPARa. У мышей с нокаутированным геном PPARa снижено пероксисомальное р-окисление липидов, подавлено образование кетоновых тел, резко снижен запас гликогена из-за торможения глюконеогенеза, что сопровождается тяжелой гипогликемией при голодании, нарушено образование жёлчных кислот из-за сниженного уровня 12а-гидроксилазы холестерина. PPARa стимулирует также со-окисление липидов за счёт индукции микросомальных оксидаз смешанного типа подсемейства CYP4A.
PPARy служит одним из факторов, определяющих дифференцировку клеток жировой ткани и, в частности — появление чувствительного к инсулину транспорта глюкозы. В дифференцированных адипоцитах PPARy увеличивает экспрессию липопротеидлипазы, транспортёров жирных кислот, ацил-КоА-синтазы, малик-фермента, фосфоенолпируват-карбоксикиназы, транспортёра глюкозы GLUT4, элементов проведения сигнала инсулина — факторов, способствующих запасанию триглицеридов. Отчасти за счёт этих эффектов через PPARy противодиабетические тиазолидиндионы восстанавливают чувствительность к инсулину при диабете типа II. Полагают, что стимулируемый отток жирных кислот в адипоциты освобождает мышцы от тормозного действия жирных кислот на утилизацию глюкозы. Выявлен ещё один механизм действия противодиабетических препаратов — торможение через PPARy образования адипоцитами TNFa и лептина, оказывающих липолитическое действие. Кроме того, снимаются тормозное действие TNFa на продукцию транспортёра глюкозы GLUT4 и фосфорилирование Рц инсулина и его субстрата в адипоцитах и тормозное действие лептина на секрецию инсулина поджелудочной железой.
Производные линолевой кислоты
Нокаут гена PPARy у мышей сопровождается гибелью эмбрионов на 10-й день из-за нарушений дифференцировки трофобласта и васкуля-ризации плаценты, истончения миокарда, а при блокировке этих неблагоприятных эффектов — гибелью в конце беременности за счёт липодистрофии и множественных кровоизлияний.
В воспалительном процессе PPARs участвуют двояким образом: провоспалительные эйкозаноиды, будучи лигандами PPARa, ускоряют собственную деградацию, а PPARy тормозит действие транскрипционных факторов (АР-1, STATs, NF-kB), стимулирующих биосинтез провоспалительных цитокинов. Показано определённое участие PPARs в атерогенезе и канцерогенезе. Предполагается, что лиганды PPARy способствуют дифференцировке моноцитов в макрофаги и далее — в пенистые клетки стенки кровеносных сосудов. Кроме того, окисленные производные линолевой кислоты в составе липопротеидов низкой плотности через активацию PPARy в резидентных макрофагах могут повышать экспрессию Рц-мусорщика CD36, усиливая тем самым отложение липидов в атеросклеротической бляшке.
PPARp, по-видимому, играет важную роль в имплантации и де-цидуализации матки. Нокаут гена циклооксигеназы-2 (СОХ2), продуцирующей природный лиганд PPARp — PGI2, нарушает эти процессы, а введение лигандов PPARp (PGI2, карбапростациклина, соединения L 165041) таким животным восстанавливает децидуализацию и имплантацию.
Rev-Erb кодируются цепью ДНК, комплементарной цепи, содержащей гены TRa и TRp. Белки Rev-Erb взаимодействуют с ДНК в форме мономеров и гомодимеров, у них отсутствует AF-2 функция, что делает их конститутивными репрессорами транскрипции. Rev-Erb служат важным элементом биологических часов, участвуют в регуляции липидного обмена. Экспрессируются часто совместно с Рц группы ROR (см. ниже) и являются их антагонистами.
Нарушение поддержания равновесия
ROR. Члены группы ядерных сиротских Рц ретиноидов ROR действуют независимо от RXR, функционируя как мономеры. RORP обнаруживается почти исключительно в элементах нервной ткани (сенсорная, нейроэндокринная и лимбическая системы). RORy экспрессируется преимущественно в скелетных мышцах, печени, почках, адипоцитах, а его сплайсинговый вариант RORy-t — в тимусе. RORa распространён значительно шире: в структурах мозга, гипофизе, адипоцитах, печени, хряще, коже, семенниках. Природный высокоаффинный лиганд ROR, по-видимому, мелатонин. Высокоэффективными лигандами ROR являются противовоспалительные препараты группы тиазолидиндионов (отличные от противодиабе-тических лекарств, действующих через PPAR). Показано участие RORy-t и RORa в иммунном ответе и стимуляции лигандами RORa образования ИЛ-2 и ИЛ-6 в мононуклеарах периферической крови. Лиганды ROR регулируют экспрессию 5-липооксигеназы — одного из ключевых ферментов биосинтеза провоспалительных лейкотриенов. При мутации или нокауте гена RORa обнаружен ряд неврологических и других отклонений от нормы (тремор, нарушение поддержания равновесия, малый размер тела, недоразвитие клеток Пуркинье мозжечка, нарушения морфогенеза в обонятельных луковицах). У таких животных значительно снижено образованиє в кишечнике аполипопротеина А-1, участвующего в транспорте липидов в форме антиатерогенных липопротеидов высокой плотности, что сопровождается склонностью к атеросклерозу. Нокаут RORb ведёт к дегенерации сетчатки и слепоте.
LXR. Рц X печени LXRa экспрессируется преимущественно в печени, а также в кишечнике, почках, селезёнке, а LXRp — повсеместно. Селективными активаторами LXR служат оксистеролы, наиболее активны из них 22(R)- и 24(8)-гидроксихолестеролы, 24(8),25-эпоксихолестерол и 7а-гидроксихолестерол.
Снижением биосинтеза жирных кислот
Оксистеролы являются промежуточными продуктами в синтезе стероидов и жёлчных кислот. У мышей с нокаутированным геном LXRa наблюдается блокировка метаболизма холестерина, его накопление в печени, нарушение регуляции гена ключевого фермента 7а-гидроксилазы (CYP7A) синтеза жёлчных кислот. В отличие от нормальных у таких животных повышение уровня холестерина не приводит к компенсаторному снижению поступления холестерина и увеличению синтеза жёлчных кислот. При низком содержании холестерина в диете существенных отклонений не наблюдается. Выключение LXRa сопровождается также снижением биосинтеза жирных кислот. В адипоцитах и перитонеальных макрофагах образуются обе формы LXR (а и (З), участвующие в регуляции обмена липидов. Нокаут гена любой из этих форм приводит лишь к частичной блокировке действия оксистеролов на экспрессию апо-Е, тогда как при двойном нокауте регуляция полностью снимается.
FXR (farnesoid X receptor — Рц X фарнезоидов) активируется высокими концентрациями фарнезола — изопренового промежуточного продукта синтеза мевалоната и далее — холестерина. Сильными индукторами FXR являются жёлчные кислоты. По-видимому, через FXR осуществляется согласование процессов повторного использования и синтеза de novo жёлчных кислот. FXR стимулирует экспрессию белка, связывающего жёлчные кислоты в кишечнике (IBABP — intestinal bile acid-binding protein), и тормозит активность промотора гена CYP7A.
PXR. Рц X прегнанов PXR экспрессируется преимущественно в печени и кишечнике. Активаторами являются антибиотик рифампицин, синтетические прегнаны, агонисты и антагонисты глюкокортикоидов и других стероидных гормонов (например, дексаметазон, прегненолон-16а-карбонитрил, антиминералокортикоид спиронолактон, антиандроген ципротерона ацетат, антиэстроген тамоксифен, фитоэстрогены). Через PXR эти лиганды стимулируют экспрессию генов семейства цитохромов Р450 ЗА, участвующих в гидроксилировании стероидов и защите от лекарств и других ксенобиотиков.
- Предыдущие статьи из рубрики: Эндокринология
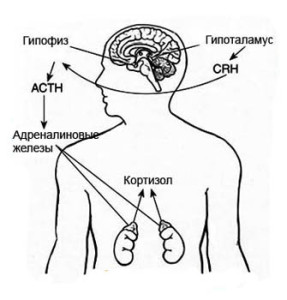 Общие свойства рецепторов
Общие свойства рецепторовРецепция и проведение сигнала Эти системы могут включать так называемые «вторые посредники» — низкомолекулярные соединения (циклические нуклеотиды, фосфолипиды […]
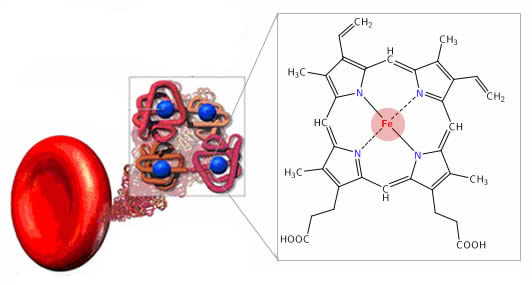 Функции транспортных белков
Функции транспортных белковОбразование эстрогенов Фермент печёночного типа (тип 1) экспрессируется в чувствительных к глюкокортикоидам тканях и преимущественно катализирует восстановительную реакцию, […]
 Транспорт сигнальных соединений
Транспорт сигнальных соединенийТранспортные белки и их лиганды К известным транспортным белкам относятся белки, связывающие стероиды (1), тиреоидные гормоны и ретиноиды […]
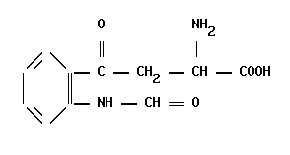 Производные триптофана
Производные триптофанаЗавершение процессов биосинтеза Так как конвертазы экспрессируются тканеспецифично, то и набор продуктов процессинга ПОМК в передней доле гипофиза, […]
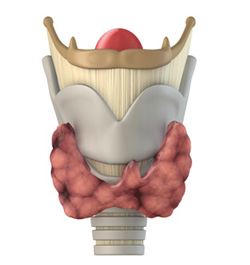 Тиреоидные гормоны
Тиреоидные гормоныЙодирование тироглобулина происходит в комплексе Гольджи у апикальной поверхности клеток под действием тиропероксидазы — гемсодержащего белка, катализирующего 1) […]