
Функции транспортных белков
Образование эстрогенов
Фермент печёночного типа (тип 1) экспрессируется в чувствительных к глюкокортикоидам тканях и преимущественно катализирует восстановительную реакцию, т.е. способствует процессам реактивации гормонально неактивных 11-кетостероидов в активные кортизол и кортикостерон. Фермент почечного типа (типа 2), обнаруживаемый в чувствительных к альдостерону тканях, преимущественно катализирует обратную реакцию, т.е. способствует инактивации глюкокортикоидов. Данный изозим играет ключевую роль в дифференцированном действии глюко- и минералокортикоидов. Известно, что Рц минералокортикоидов одинаково хорошо взаимодействует с альдостероном и глюкокортикоидами. Учитывая, что концентрация глюкокортикоидов в крови на 2—3 порядка превышает концентрацию альдостерона, в отсутствие механизма дискриминации глюкокортикоидов альдостерон не мог бы выполнять своей специфической регуляторной функции. Функцию такого дискриминатора выполняет Пр-ГСД типа 2, инактивирующая глюкокортикоиды, но не влияющая на альдостерон, 1 ір-гидроксильная группа которого недоступна для фермента из-за образования 11—18 гемиацеталя.
Превращение андрогенов в эстрогены под действием ароматазы в периферических тканях — минорный путь метаболизма андрогенов, но он служит важным источником эстрогенов в организме. Ароматаза экспрессируется во многих тканях, включая мозг, жировую ткань, мышцы. У грызунов перинатальная необратимая индукция ароматазы под действием андрогенов семенников в ряде областей мозга в значительной мере обеспечивает последующую половую дифференцировку нейроэндокринной системы и поведения. Образование эстрогенов в жировой ткани сглаживает переход к менопаузе у тучных женщин. Следует отметить, что 5а-дигидротестосте-рон является неароматизируемым андрогеном, и его применяют для выяснения эндокринного механизма действия тестостерона (прямого или через трансформацию в эстрогены).
Предовуляторный период
Наличие в периферических тканях Зр-олдегидрогеназы обеспечивает возможность активации секретируемого в значительных количествах надпочечниками дегидроэпиандростерона (DHEA) в цепочке превращений DHEA -» андростендион —> тестостерон —> 5а-дигидро-тестостерон.

Стероидные гормоны могут модифицироваться также путём дополнительного гидроксилирования под действием ряда цитохромов Р450. Данные превращения ведут к снижению или утрате гормональной активности стероидов, но, как правило, представляют минорный путь метаболизма. В то же время образование в гипоталамусе катехолэстрогенов при гидроксилировании эстрогенов в положениях 2 или 4 играет важную роль в системе положительной обратной связи в предовуляторный период. С одной стороны, кате-холэстрогены, обладая очень слабой эстрогенной активностью, но будучи эффективными лигандами Рц эстрогенов, блокируют систему отрицательной обратной связи эстрогенов на продукцию го-надолиберина. С другой стороны, катехолэстрогены эффективно конкурируют с индукторами секреции гонадолиберина катехолами-нами за фермент катаболизма катехоламинов — катехол-О-метил-трансферазу. В результате эффективность стимулирующего действия катехоламинов на гонадолиберин возрастает. Катехолэстрогены через образование соответствующих хинонов способны образовывать аддукты ДНК, что может иметь отношение к известному проканцерогенному действию эстрогенов. Образование катехолэстрогенов кататизируют цитохромы Р450 ряда семейств (CYP1A1, CYP1A2, CYP1B1, CYP2C9, CYP3A4). В условиях индукции цитохромов Р450 (под действием лекарственных препаратов или поллютантов) гид-роксилирование стероидов может становиться количественно сопоставимым с рассмотренными выше путями обмена. Цитохромы Р450 не обладают строгой региональной субстратной специфичностью. Например, CYP1B1 катализирует гидроксилирование эстрогенов в положениях 2, 4 и 16а.
Образование парных соединений
Выведение продуктов обмена стероидов из организма облегчается образованием парных соединений с глюкуроновой и серной кислотой. Реакции сульфирования катализируются ферментами семейства сульфотрансфераз, использующих в качестве второго субстрата активированную форму серной кислоты — З’-фосфоадено-зин-5′-фосфосульфат (PAPS).
Сульфотрансферазы, подразделяемые на 6 подсемейств, обладают ограниченной субстратной специфичностью. Сульфирование эстрогенов преимущественно катализируют эстрогенсульфотрансферы (подсемейство SULT1E), а также некоторые фенолсульфотрансферазы (подсемейство SULT1A). Сульфирование 3-гидроксистероидов (метаболитов кортикостероидов, прогестинов, андрогенов), а также жёлчных кислот преимущественно катализируют гидроксистероид-сульфотрансферазы (подсемейства SULT2A и SULT2B). У грызунов экспрессия сульфотрансфераз, активных на эстрогенах и андрогенах, дифференцирована по полу с преобладанием у самцов и самок соответственно, что, по-видимому, служит защитой от избытка гормонов противоположного пола. Нокаут гена эстрогенсульфотрансферазы у мыши ведёт к нарушению структуры и функций семенников. Образование сульфатов может также служить формой депонирования гормонов в организме, а дегидроэпиандростеронсульфат проявляет активность антагониста Рц ГАМ К типа А.
Присоединение глюкуроновой кислоты к гидроксильным группам стероидов происходит под действием глюкуронозилтрансфераз, использующих в качестве активированного второго субстрата УДФ-глю-куронат.
Экспрессия ферментов семейства
Клонировано 15 изозимов фермента человека, которые делят на 2 подсемейства. Представители обоих семейств способны катализировать образование парных соединений стероидов. Строгой субстратной специфичности ферментов нет, хотя имеется определённая региональная специфичность. Например, изозим УДФ-глюкуронозилтрансферазы UGT1A1 (UDP-glucuronosyltransferase) катализирует присоединение глюкуроновой кислоты по положению 3 молекулы эстрогенов, а изозим UGT2B17 катализирует присоединение по 3 и 17 положениям. Помимо стероидов, УДФ-глюкуронозил-трансферазы используют в качестве субстратов множество других эндогенных соединений (например, билирубин, тиреоидные гормоны) и ксенобиотиков, участвуя в их детоксикации. Ферменты данного семейства (например, UGT2B7) способны также катализировать гликозилирование субстратов (в частности, жёлчных кислот) с использованием УДФ-глюкозы в качестве активированного второго субстрата. Экспрессия ферментов семейства дифференцированно регулируется гормональными и другими факторами. В частности, в клетках простаты андрогены снижают активность изозима UGT2B17, но не UGT2B15, инактивирующих эти гормоны.
Образование парных соединений стероидов происходит преимущественно в печени, но может иметь место также в местах их синтеза (гонадах, коре надпочечников) и тканях-мишенях (молочной железе, матке, предстательной железе), а также в почках, коже, кишечнике.
Сульфаты и глюкурониды стероидов могут быть гидролизованы с высвобождением исходных стероидов под действием соответствующих ферментов. Десульфирование катализируется стероидсульфатазой (арилсульфатазой С). Делеция или точечные мутации гена этого фермента сопровождаются развитием одной из форм ихтиоза в виде отложения в коже субстрата фермента — сульфата холестерина. Активность этого фермента в молочной железе и её опухолях служит важнейшим источником поступления активных эстрогенов в ткань.
Гидролиз глюкуронидов
Гидролиз глюкуронидов катализирует р-глюкуронидаза. Фермент экспрессируется в ряде тканей, включая простату, почки, кишечник, а также кишечной флорой. В почках активность фермента контролируется половыми гормонами, гормоном роста, инсулино-подобным фактором роста I. Фермент широко используется для анализа метаболитов различных соединений, включая стероидные анаболики, в моче и кале.
Метаболизм основного гормонально активного ретиноида — трансретиноевой кислоты — включает обратимую изомеризацию в 9- и 13-цис-ретиноевые кислоты, образование парного соединения с глюкуроновой кислотой, гидроксилирование в положениях 4 и 18, образование 5,6-эпоксида и введение дополнительной двойной связи в кольцевую часть молекулы.
4- и 18-Гидроксилирование ретиноевой кислоты и её изомеров может катализироваться рядом цитохромов Р450 (CYP2C8, CYP2C9, CYP3A4, CYP3A7, CYP26A1). Особую роль, по-видимому, играет CYP26A1, поскольку этот фермент, во-первых, высокоспецифичен в отношении ретиноидов и, во-вторых, индуцируется ретиноевой кислотой, обеспечивая, очевидно, систему ауторегуляции уровня ретиноевой кислоты. Эпоксидирование транс-ретиноевой кислоты и её изомеров катализирует простагландин-Н-синтаза. Реакцию потенцирует арахидоновая кислота и другие соединения, служащие субстратами для образования перекисей. Метаболические превращения ретиноидов происходят в присутствии клеточных белков, связывающих ретиноевую кислоту [CRABP (cellular retinoic acid binding protein) 1 и 2], относящихся к семейству липидсвязывающих белков. Эти белки ограничивают доступность лигандов для Рц ретиноидов, но не препятствуют метаболизму, поскольку их комплексы с ретиноидами узнаются рядом метаболизирующих ферментов как субстраты с величинами Кт даже более низкими, чем для свободных субстратов. CRABP-2, по-видимому, участвует в системе ауторегуляции действия ретиноидов, поскольку его экспрессию индуцирует ретиноевая кислота.
Парные соединения с глюкуроновой кислотой
Сама ретиноевая кислота и многие её метаболиты могут образовывать парные соединения с глюкуроновой кислотой. Остаток глюкуроновой кислоты может присоединяться с помощью УДФ-глю-куронозилтрансфераз к карбоксильной группе ретиноевой кислоты и её 4-оксо- и 5,6-эпоксипроизводных, а также к гидроксильной группе 4-гидроксиретиноевой кислоты. Ведущую роль в этих процессах у человека играет изозим UGT2B7. Окисленные продукты ретиноевой кислоты сохраняют гормональные свойства родительского соединения, хотя и в ослабленной (в 2,5-6 раз) форме. Образование глюкуронидов, по-видимому, дополнительно снижает активность ретиноидов и облегчает их выведение из организма с мочой и/или жёлчью.
Эйкозаноиды. Инактивация простагландинов (PGs) начинается с окисления общей для всех PGs гидроксильной группы в положении 15(S) до кетогруппы, катализируемого 15-простагландиндегид-рогеназой (семейство дегидрогеназ/редуктаз с короткой цепью). Данный фермент участвует, в частности, в защите жёлтого тела от индуцируемого PGF2a лизиса во время лютеиновой фазы цикла и предотвращает преждевременные роды. В конце беременности глюкокортикоиды плода ингибируют экспрессию фермента, что способствует инициации родовой деятельности. Следующей стадией катаболизма PGs служит восстановление имеющейся во всех PGs двойной связи в положении 13, катализируемое д-13-15-кетопро-стагландинредуктазой. Указанный фермент обладает также лейко-триен В4 12-гидроксидегидрогеназной активностью. Окисление тромбоксана ТХВ2 в положении 11 с образованием 11-дегидро-ТХВ2 катализирует 11-гидрокситромбоксан В2-дегидрогеназа. Эйкозаноиды могут также катаболизироваться путём омега-гидроксилирова-ния, катализируемого цитохромами Р450 (подсемейства CYP4A и CYP4F), с образованием 18- и 19-гидроксипроизводных. Указанные ферменты проявляют определённую предпочтительность в отношении PGs или лейкотриенов и дифференцированно экспрессируются в полиморфноядерных лейкоцитах (CYP4F3), семенных пузырьках (CYP4F12), печени и многих других тканях (CYP4F2, CYP4F3B).
- Предыдущие статьи из рубрики: Эндокринология
 Транспорт сигнальных соединений
Транспорт сигнальных соединенийТранспортные белки и их лиганды К известным транспортным белкам относятся белки, связывающие стероиды (1), тиреоидные гормоны и ретиноиды […]
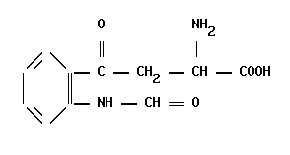 Производные триптофана
Производные триптофанаЗавершение процессов биосинтеза Так как конвертазы экспрессируются тканеспецифично, то и набор продуктов процессинга ПОМК в передней доле гипофиза, […]
 Тиреоидные гормоны
Тиреоидные гормоныЙодирование тироглобулина происходит в комплексе Гольджи у апикальной поверхности клеток под действием тиропероксидазы — гемсодержащего белка, катализирующего 1) […]
 Производные аминокислот
Производные аминокислотФосфолипазы А2 — большое семейство (известно около 20 членов) белков с различающейся субстратной предпочтительностью, локализацией (выделяют секретируемые, зависимые […]
- Биосинтез фосфолипидов
Биосинтез фосфолипидов группы РАР, обладающих сигнальными свойствами, может идти двумя путями. При биосинтезе de novo (преобладающем в почках […]